|
|
|||
|
БІОСИСТЕМИ. БІОСФЕРА
ЕКОСИСТЕМИ. ПОПУЛЯЦІЇ
ОРГАНІЗМИ У ДОВКІЛЛІ
ЛЮДСТВО ТА ЙОГО ДОЛЯ
|
|||
|
← IV-17. (доповнення) Як паразити «підставляють» своїх хазяїв |
IV-18. (доповнення) Стратегії внутрішньопопуляційноі взаємодії |
||

IV-18. (доповнення) Стратегії внутрішньопопуляційної взаємодії
Можна сказати, що значна частина організмів живе у двох різних за своєю суттю середовищах: зовнішньому та внутрішньопопуляційному. Внутрішньопопуляційне середовище — це сукупність відносин між родичами, які не вичерпуються наведеними нижче прикладами.
Одним із найважливіших механізмів внутрішньо популяційного регулювання чисельності є територіальність — конкуренція між особинами популяції за використання простору з усіма його ресурсами. Часто територіальність проявляється у захисті індивідуальної території. Найчастіше площа цієї території більша, ніж мінімально необхідна для виживання особини чи її сім'ї. У різних організмів територіальність проявляється по-різному. Іноді індивідуальні ділянки охороняють лише самці, іноді пари, іноді як самці, так і самки конкурують одне з одним за індивідуальні ділянки.
Ви замислювались, навіщо співають співочі птахи? На етапі утворення пар самці співають, щоби привабити самок. Втім зрештою пари утворені, гнізда побудовані, з яєць вивелися пташенята, а самці все ж продовжують співати... На цьому етапі спів самців показує, що сімейна ділянка зайнята. Як не дивно, за своїм біологічним змістом ця поведінка досить близька до поведінки домашнього собаки, який на іншому кінці повідця під час прогулянки з господарем мітить своєю сечею кожен кут будинку, стовп чи дерево. У більшості випадків територіальні особини знають, де закінчується їхня ділянка, а де починаються ділянки сусідів. Положення кордону — результат урівноваження зусиль господарів сусідніх територій щодо розширення своїх ділянок. Порушення чужої території викликає сутички, причому результат цих сутичок зазвичай залежить від цього, на чиїй території вони відбуваються.
«Комплекс територіальної поведінки не вичерпується прямими нападами, сутичками, перегонами тощо. Щобільше, такі жорсткі форми охорони в чистому вигляді трапляються досить рідко; майже завжди агресія супроводжується ритуальними формами поведінки: пози загрози, специфічні звукові сигнали, демонстрація нападу без доведення його до фізичного контакту тощо. Значення загрози має, наприклад, певний нахил тулуба до горизонталі у ряду видів риб, «підвищені» пози деяких безхвостих амфібій, «кивання» головою, підіймання та опускання передньої частини тіла та розправлення горлової складки у ящірок-ігуанід, різноманітні форми демонстрації у різних ссавців. Здебільшого демонстрації загрози сприймаються особиною, яка потрапила на чужу ділянку, як сигнал до втечі» (І.О. Шилов, 1998).
Найчастіше в територіальній популяції тварин розмножуються не всі особини, а лише ті, хто має індивідуальну ділянку. «Волоцюги» змушені залишатися бездітними, чекаючи випадку, коли їм вдасться зайняти територію, що звільнилася в результаті дій хижака або відбити ділянку у господаря території, що постарів. Така поведінка є ефективним способом стабілізації чисельності популяції: у будь-який момент розмножується стільки особин, скільки на цій території існує індивідуальних ділянок. Втрати поголів'я господарів ділянок не є критичними: на їх місце тут же заступлять особини з числа «волоцюг», популяційного резерву. Територіальність — не єдиний внутрішньо популяційний механізм, що обмежує зростання чисельності популяції. Наприклад, у мишоподібних гризунів регуляція плодючості залежно від щільності здійснюється гормональним шляхом. При надмірній щільності популяції кількість контактів між родичами перевищує певну межу та викликає реакцію стресу та гормональні перебудови, що ведуть до зниження плодючості.
З попереднього викладу могло скластися враження, що популяція — це арена боротьби та безкомпромісної конкуренції всіх проти всіх. Звісно, це не так. Біологія знає безліч прикладів взаємодопомоги та підтримки у популяції. Особливо цікаво зрозуміти причини альтруїстичної поведінки, тобто такої, коли одна особина завдає собі якоїсь шкоди або навіть жертвує життям заради інших. Найлегше зрозуміти таку поведінку, коли йдеться, наприклад, про матір, яка захищає свій виводок. Навіть якщо мати гине, можлива ситуація, коли сумарна репродуктивна цінність (тобто цінність майбутнього популяції) потомства вища за цінність самої матері. Наприклад, якби існував ген, який змушує матір жертвувати своїм життям заради дітей, такий ген міг би поширюватися в популяції завдяки кращій виживаності дітей альтруїстичних матерів.
Наведене міркування цілком відповідає духу соціобіології — науки, яка пояснює поведінку тварин (і, зокрема, людини) як результат добору генетично визначених ознак. Творцем соціобіології вважається американський фахівець з поведінки комах Едуард Вілсон, який опублікував 1975 року книгу «Соціобіологія. Новий синтез». Логіку цієї науки найпростіше показати на прикладі спорідненого добору, описаного одним із класиків соціобіології, Вільямом Гамільтоном. Гамільтон зміг пояснити, чому найчастіше альтруїстична поведінка реєструється у соціальних перетинчастокрилих — бджіл, ос і мурах. Річ у тому, що для цих комах характерна досить незвична система хромосомного визначення статі. Самка цих комах може відкладати незапліднені яйця (з одним хромосомним набором), з яких виходять самці, та запліднені (з двома), з яких виводяться самки. Зазвичай подібність генів батьків та дітей, і навіть дітей певних батьків одного з одним становить 1/2 (половина генів подібні, половина — ні). Однак у перетинчастокрилих ситуація інша. Подібність матері та її дочок — 1/2, а ось сестер одна з одною — 3/4 (оскільки спадковість батька складається з єдиного набору хромосом, усі дочки одержують його повністю). Це означає, що самка забезпечить краще виживання власних генів, якщо доглядатиме за своїми сестрами, а не за своїми дочками! Очевидно, ці механізми справді є причиною виникнення сімей соціальних комах, де розмножується одна самка (матка чи цариця), а більшість особин представлена безплідними самками, які доглядають за потомством матки — за власними сестрами. Описаний механізм, що зумовлює альтруїстичну поведінку, діє лише на самок. І справді, наприклад, у бджіл самці (трутні) не беруть участі у спільній діяльності сім'ї.
Втім, соціальність у комах можлива не тільки на основі описаного генетичного механізму. Доказом цього є терміти. Ймовірно, причина їхньої соціальності інша — колективне травлення, при якому для розщеплення целюлози особини змушені багато разів їсти екскременти один одного. До речі, у термітів і самки, і самці роблять однаковий внесок у «громадські» роботи.
Чудово пояснюючи багато дивовижних особливостей поведінки комах, соціобіологія «спрацьовує» трохи гірше, коли йдеться, наприклад, про ссавців. Це може бути пов'язано з багатьма причинами, зокрема з тим, що у тварин з гнучкою поведінкою, певно, відсутні гени, які жорстко зумовлюють той чи інший спосіб дій у певній ситуації. Придатність висновків соціобіології до опису поведінки людини досі залишається предметом запеклих наукових та навколонаукових суперечок.
Наприклад, майже все потомство в стаді павіанів належить кільком домінантам (вожакам), які, об'єднуючи свої зусилля, утримують самців-субдомінантів у підлеглому стані (зокрема, забороняючи тим самим паруватися із самками). Періодично, в міру старіння ватажків, у групі павіанів відбуваються «революції»: субдомінанти спільними зусиллями «скидають» старих ватажків та отримують пріоритетний доступ до їжі та самок. Однак у разі зовнішньої небезпеки (наприклад, напад леопарда) і домінанти, і субдомінанти об'єднують свої оборонні зусилля і часто жертвують своїм життям, захищаючи групу (рис. IV-18.1). З погляду соціобіології молодий самець, який віддає життя за групу до того, як він залишив потомство, чинить «неправильно». Однак якщо ми розглянемо цю ситуацію не з погляду добору окремих генів, а з погляду особливостей психіки самця як ватажка, який не досяг статусу лідера, нам стане зрозуміла готовність мавпи відганяти хижака від зграї за будь-яку ціну.
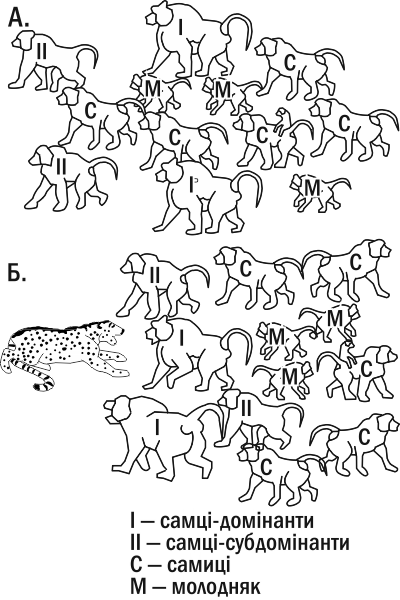
Рис. IV-18.1. У павіанів зареєстрована здатність бездітних самців йти на самопожертву, захищаючи зграю. Тут спрощено показаний похідний порядок павіанів (А.) та їх лад при нападі хижака (Б.)